Introduction
The brachiocephalic muscle connects the brachium to the skull or cranial cervical region. This muscle is relatively complex than other muscles in domestic animals because the clavicle should be reduced into fibrous intersection, called the clavicular intersection, which results in the fusion between the cleidomastoid muscle and the clavicular part of the deltoid muscle. As a result, in domestic mammals, the brachiocephalic muscle consists of two or three parts. These portions are named by their relationship to origin, clavicular intersection. From the origin the caudal part of the muscle, the cleidobrachial muscle, inserts at the humerus in all domestic mammals. However, its cranial attachments appear to vary among species of domestic mammals. For example, in carnivores they consist of the cervical part and the mastoid part of the cleidocephalic muscle [1-10]. In ruminants and pigs, the attachments consist of the mastoid part of the cleidocephalic muscle and the occipital part of the cleidocephalic muscle and in horses, they consist of the mastoid part of the cleidocephalic muscle [5-10].
Recently, several reports on variation in skeletal muscles, such as the hyoid muscles and the neck muscle in dogs, have been published [12-18]. In veterinary anatomical practice the variations and complexity in the skeletal muscles may make it difficult for the students to learn about the structures of neck. In addition this complexity could confuse veterinarians and complicate surgical procedures in this area. Therefore, information about its normal structure and variation therein is necessary. Herein, we provide the second reported case of a variation of the brachiocephalic muscle in a dog.
Case report
During routine dissections of a cadaver of an adult mixed-breed male dog of approximately 12.0 kg body weight, we noted a variation in the insertion of the mastoid part of the cleidocephalic muscle in left and right. The brachiocephalic muscle was made of four bellies, which were the cleidobrachial muscle, the cervical part of the cleidocephalic muscle, and the two bellies of the mastoid part of the cleidocephalic muscle. Their origin, the clavicular intersection in front of the shoulder joint, was about 3.2 cm in length. The cleidobrachial muscle was 8.5 cm in length and 0.1 cm in thickness. The cervical part of the cleidocephalic muscle was 2.5 cm wide and 0.1 cm thick and cranially attached by an aponeurosis of 10.0 cm wide on the dorsal fibrous raphe for the cranial half of the neck. The mastoid part of the cleidocephalic muscle was composed of two bellies, inserted independently at the mastoid process of the temporal bone and the ventral surface of the wing of the atlas (Fig. 1). This portion was covered by the cervical part of the cleidocephalic muscle and the occipital part of the sternocephalic muscle. We could conveniently call the division attached to the atlas the dorsal belly and that attached to the temporal bone the ventral belly because they were situated dorsally and ventrally and then ran cranially together, although a portion of their origin overlapped. The ventral belly extended from the clavicular intersection and was 0.9 cm wide, 0.4 cm thick, and 12.5 cm long. It inserted by means of a thick tendon at the mastoid process of the temporal bone with the mastoid part of the sternocephalic muscle that lies ventral to it. The dorsal belly of the mastoid part of the cleidocephalic muscle started from the clavicular intersection and was 1.0 cm wide, 0.2 cm thick, and 11.5 cm long. The latter narrowed as it coursed cranially and inserted at the wing, being 0.4 cm wide. The mastoid part of the cleidocephalic muscle was innervated by the accessory nerve and the ventral branches of the second, third, and fifth cervical nerves and was supplied by the ascending branch of the superficial cervical artery. Other variations or abnormalities were not found.
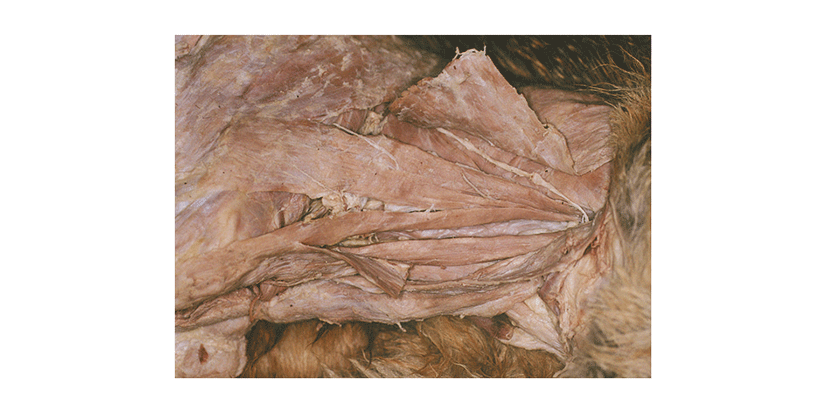
Discussion
Knowledge of muscle variation in dogs is essential in both the field of veterinary anatomy and in animal clinics because the animals are the most frequent subjects used for teaching as well as the most common patients of domestic veterinarians. Recently, attention has been paid to variation in skeletal muscles. Among hyoid and thyroid muscles, variation has been reported in the stylohyoid, sternohyoid [13, 14, 16], mylohyoid [14], sternothyroid [15], and thyrohyoid [18]. Among leg muscles, variation has been reported in the soleus muscle [19] and the extensor digitorum brevis muscle [20]. Recently, Alić et al [12, 13] reported variation in pectoral girdle muscles and neck muscles including the brachiocephalic, sternocephalic, omotransverse, capital rhomboid, cervical ventral serrate, and dorsal scalenus muscles. Terrado et al [17] reported a case of variation in the splenius muscle in a dog. In most of these cases, there was an additional belly that inserted at the same point as a normal belly or there was an anomaly attached to other sites.
The action of the brachiocephalic muscle in the dog is to draw the limb cranially, to fix the neck acting bilaterally or to fix the head and neck acting unilaterally. The mastoid and the cervical parts of the cleidocephalic muscle are innervated by an accessory nerve and the ventral branches of the cervical nerves and the cleidobrachial muscle by the ventral branches of the sixth and seventh cervical nerves. The cleidocephalic muscle reaches a width of 2.5 to 3 cm and a thickness of 7 to 10 mm in a medium-sized dog and it is often split into two round bundles throughout its length [2, 3, 8].
In the present case, the mastoid part of the cleidocephalic muscle consisted of two bellies with different insertions. The ventral belly inserted normally at the mastoid process of the temporal bone, but it was half the normal size for a medium-sized dog [3]. The dorsal belly was attached to the atlas, but was innervated by the same nerve and supplied by the same artery as the belly ventral to it. However, in combination, the two bellies were similar in size to the one belly of a normal medium-sized dog. Therefore, we presumed that these two bellies were of the same origin. Our case is different from that report by Alić et al [12]. In that case, the cranial end of mastoid part of the left cleidocephalic muscle was divided into two parts and inserted at the mastoid process of the temporal bone and the nuchal crest, respectively.
The cause of that variation was not declared in case studies; the relationships of muscles in phylogenetic changes from reptile to mammals is only conjecture [14, 15]. For example, the clavicle and coracoid bone are well developed in reptile, bird, and monotrems [8, 11]. These bones play a role in the attachment sites of neck muscles. However, these bones are reduced or fused in domestic mammals because the relationship of limbs with body had to be changed. As results, the muscles could be attached on adjacent structures. Another possibility of muscle variation conjectured is a migration problem of the myotome during early embryonic development [12, 18]. The authors presumed either that the myotomal cells had not differentiated or that they died for unknown reason during the developmental stage in case of absence of the muscle. If the myotomal cells might have differentiated but migrated to a near location, a variation in muscle attachment should occur.
Recently, Alić et al [12, 13] surveyed muscle variation in the neck and pectoral girdle of 57 cross-bred dogs and reported frequencies of 7.0% in the neck and 12.3% in the pectoral girdle. These percentages are higher than expected. Therefore, such variations in dogs may more common than previously assumed and should be considered during anatomical dissections and surgical procedures.